Evidence of diversification through geographic isolation
|
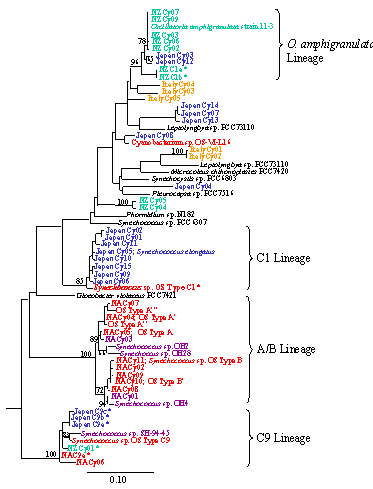
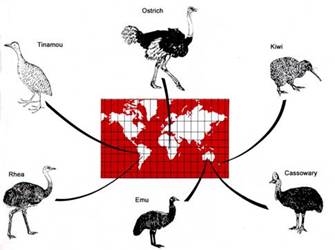
Since bacterial diversity seems to arise through adaptive radiation, it seemed of interest to examine whether the other major driver of speciation in plants and animals, geographic isolation, was important to prokaryote diversification. Considering hot springs to be like islands, we sampled springs in Japan, New Zealand and Italy in addition to North America. We retrieved sequence data for both the 16S rRNA and ITS loci of indigenous cyanobacteria (Fig. 6) directly from the mats we sampled. We found unique cyanobacterial 16S rRNA genotypes in each location, as shown in Fig. 7. The A/B typeSynechococcus sequences appear endemic to North America (red and pink for OR). Japan (blue) was dominated by C1-lineage Synechococcus, which were also found in North America. New Zealand (green) was dominated by C9-typeSynechococcus and Oscillatoria amphigranulata sequences. In many cases we observed distinct clades for different geographical regions. The geographic distribution pattern could not be explained by different chemical conditions, suggesting that geographic isolation is involved in diversification of hot spring cyanobacteria. We even found evidence of geographic clades at more local spatial scale (e.g., within major thermal basins of Yellowstone Park. Together with similar results from other laboratories, it appears clear that, like plants and animals (Fig. 8), geographic isolation also acts upon prokaryote variation to cause diversification. [Papke et al., 2003] |
Figure 7. Cyanobacterial 16s rRNA diversity detected in globally separated hot spring mats of North America (OR),Japan, and New Zealand. A/B, C1, and C9 lineages areSynechococcus spp. |
||
|
Fig. 8. Distinct flightless birds on different continents exemplify diversification through geographic isolation. [Begon et al., 1990] |